작물생리학 핵심 요점 정리 농업직 공무원, 농촌지도사, 농업연구사 시험대비 6. 작물의 발아와 휴면생리
6. 작물의 발아와 휴면생리
* 종자발아 순서 : 수분흡수 → 효소활성화, 저장양분 소화 → 양분 분해 및 이동 → 호흡 → 유식물의 생장, 발육
☞ 흡수 ⇨ 양분 이화 ⇨ 이동 ⇨ 산화 ☞ 발아과정의 생리상 : 흡수기(A相), 발아준비기(수분포화기 ; B相), 생장기(C相)

☞ 가수분해 효소 활성화(지베렐린) ⇨ 세포분열 촉진 / 유근, 유아 생장(시토키닌) ⇨ 세포신장 촉진, 뿌리 지하로, 어린싹 지상(옥신)
☞ 단자엽 식물의 경우 배에서 생성된 지베렐린은 배반을 통하여 방출되어 호분층으로 이동
* 배의 생장
① 배를 구성하고 있는 세포들의 신장
② 유아와 유근을 연결하는 역할을 맡은 배축의 형성과 생장
③ 유아나 유근의 분열조직에서 새로운 세포의 생성에 따른 형태적 변화과정을 통해 유식물체 형성

* 논벼의 저온발아성 정도 : 도입 일본종 < 국내육성종 < 재래종 / 조만성 : 만생종 < 중생종 < 조생종
↳ 논벼 종자에 대한 GA3(100ppm), 에틸렌(5~10ppm), r선 등의 처리는 저온발아성 향상
* 산소와 발아
- 수중 불발아 종자 : 밀, 귀리, 무, 배추, 가지, 고추, 알팔파, 양배추, 코스모스, 후추, 파, 메밀, 콩, 알팔파
- 수중 발아 저하 종자 : 미모사, 담배, 토마토, 화이트클로버, 카네이션, 석죽
- 수중 발아 종자 : 벼, 상추, 당근, 셀러리, 피튜니아, 티머시, 켄터키블루그래스
* 광(光)과 발아
- 호광성 종자(목초, 화본과) : 화본과 목초, 담배, 상추, 우엉, 셀러리, 갓, 뽕나무, 페튜니아, 베고니아
- 수중 발아 저하 종자 : 미모사, 담배, 토마토, 화이트클로버, 카네이션, 석죽
- 수중 발아 종자 : 벼, 상추, 당근, 셀러리, 피튜니아, 티머시, 켄터키블루그래스

※ 광 파장은 적색 부분(660~700nm)으로 그중 670nm에서 가장 촉진되고, 700nm에서 억제
* 이유기와 독립생장기의 전환
유식물은 초기에 배유 자엽의 저장양분에 의해서 생육, 독립영양의 시기로 변천하게 되는데 이 전환기를 이유기라고 한다.
→ 배유양분 소진기 생장상태 : 벼 본엽 4매, 밀 본엽 3매, 보리 본엽 2매
* 발아 촉진 물질
| 지베렐린 | GA3가 많이 이용, 발아에 필요한 광과 온도를 대체할 수 있는 효과 - 광발아종자의 발아 촉진 |
| 시토키닌 | 정아우세를 억제하고 측아의 생장을 촉진, 광선과 결합하여 발아를 촉진하고, 키네틴은 호광성 종자의 암발아를 유도한다. - ABA의 작용을 억제하므로 발아 촉진 |
| 에틸렌(C2H4) | 과실의 성숙이나 눈의 휴면 또는 잎의 이층 형성에 효과, 종자의 발아를 촉진 ☞ 상추, 양딸기, 땅콩, 유채, 서브클로버 종자의 발아촉진 |
| 과산화수소(H2O2) | 침엽수, 콩과식물, 토마토, 보리 등의 발아 촉진, 종자 살균제 역할도 가능 |
| 질산칼륨(KNO3) | 발아 촉진에 쓰이며 발아시험에는 0.1~1.0%의 농도로 이용, 화본과 목초의 발아촉진, 암중발아 촉진 |
| Thio - Urea | 발아 촉진에 널리 이용, 발아에 필요한 광과 온도를 대체 할 수 있는 효과 |
| MnSO4 | 상당히 넓은 농도범위에서 망고, 옥수수, 양배추 등의 종자발아 촉진 |
* 발아 촉진 / 휴면타파방법

습윤저온처리(층적법)시 휴면종자의 후숙이 진행되는 동안 생리적 변화(0℃ 이하에서는 효과 없고 5℃ 내외에서 효과 크다)
① lipase, peroxidase, oxidase, catalase 등 효소활력의 증가 ② 삼투퍼텐셜 저하
③ 불용성 물질이 가용성 물질로 변화 ④ 신조직 형성에 사용되는 당류, 아미노산 등의 집적
* 휴면타파
① 미숙배의 휴면타파 : 층적법(저온 습윤 처리) → ABA, Coumarin 저하 / GA3, cytokinin 상승
② 화학물질 처리에 의한 휴면타파 : thiourea, ethylene chlorohydrin, dinitrophenol, NaN3(sodium azide),
NaC7H5O3(sodium salicylate), CO2, N2, ethylene
③ 식물호르몬에 의한 휴면타파 : GA, Cytokinin은 함께 있으면 종자의 휴면타파하고 촉진되나,
Cytokinin은 혼자 있으면 억제물질의 존재유무와 관계없이 휴면타파를 하지 못한다.
⇨ 발아조건 : ① GA ○, Cytokinin ○, ABA ○ / ② GA ○, Cytokinin ○, ABA × / ③ GA ○, Cytokinin ×, ABA ×
☞ GA는 발아에 있어 필수적으로 필요하며 Cytokinin은 GA와 함께 존재해야만 발아 할수 있으며 GA와 ABA가 동시에 존재한다면 휴면하게 된다.
* 종자의 발아력 상실
① 원형질 단백질 응고 ② 효소활력 저하 ③ 생리활성물질 활력저하 ④ 양분소모 ⑤ 종자에 수분 함량 저하
* 발아 억제 물질 : 종자나 과피의 껍질에 존재하는 발아 억제 물질에는 암모니아, 시안화수소, 알데하이드, 쿠마린, 페놀산, 앱시스산 등이 있다.
☞ HCN, H2S 등은 발아율에 별로 영향 주지 않으나 NH3, Cl 등은 발아에 유해, 특히 SO2의 해가 뚜렷
☞ 감자의 저장중 발아억제제 : NAA, MENA - 맹아억제
☞ 밤종자의 저장에 2,4-D, NAA가 효과적 이용, NAA는 밀의 수발아 억제에도 효과가 있다.
* 종자 휴면의 유형 : ① 불투수성 종피 ② 종피의 기계적 저항에 의한 휴면 ③ 산소를 통과시키지 않는 종피
④ 미발달된 배에 의한 휴면 ⑤ 2차 휴면
불투수성 종피 : 경실 종자 - 물이 종피를 투과하지 못하여 장기간에 걸쳐 발아 할 수 없는 종자
↳ 콩, 팥, 동부, 감자, 오크라, 황마, 나팔꽃, 연, 붉은토끼풀, 흰토끼풀, 갸자리
- 루핀, 헤어리베치 등의 종자는 고온, 저습 저장시 경실화 / 수분의 상태에 따라 경실 – 비경실로 구분 가능
- 종피의 불투수성은 울타리세포(책상세포)와 관련
※ 울타리 세포안의 pectin이 불투수성에 관여하는 물질로 알려져 있다.
펙틴 함량이 많을수록 경실화 되는 경향이 많으며 울타리 세포안의 suberin이 불투수성에 관여한다.
☞ 종피의 불투수층은 책상조직이며 관여물질은 책상세포 안의 펙틴(pectin)이 지적되고 있다.
* 경실종자의 휴면타파법
① 파상법 : 종피에 기계적 상처를 만들어 흡수시키는 방법
② 화학적파상법 : 화학물질 (진한황산, 수산용액 등)로 종피에 상처를 만들어 흡수시키는 방법
* 종피의 불투수성 원인 : ① 건조에 의한 종피의 기계적인 수축 ② 토양수분이 많을수록 ③ 소립종자일수록
④ 숙도가 높은 종자일수록 ⑤ 저장에 의하여 종자가 경실화

* 종자의 수명 ☞ 함수량이 적을 때 종자는 비교적 저온에 잘 견디며, 어느 정도까지 함수량을 줄이면 액체질소의 온도에서도 해를 받지 않는다
| 단명종자 (3년 이내) microbiotic seed | 콩, 양파, 파, 당근, 땅콩, 고추, 상추, 메밀, 우엉, 토당귀 등 |
| 상명종자 (3 ~ 15년) mesobiotic seed | 벼, 보리, 밀, 무, 완두, 목화 |
| 장명종자 (15 ~ 100년) macrobiotic seed | 토마토, 수박, 호박, 녹두, 오이, 배추, 가지, 알팔파 |
☞ 종자의 수명에 영향을 끼치는 외적조건 : ① 습도 ② 온도 ③ 산소
☞ 함수량이 많은 종자를 완전히 산소가 제거된 장소 보관시 혐기성 호흡으로 유해물질 생성되어 발아저해,
장기저장에서 산소의 존재가 수명을 단축하는 결과 초래
* 종자의 수명에 미치는 영향
① 일반적으로 건조종자의 수명은 수분을 다량 함유한 종자보다 길다. 이는 함수량이 많을 때 단백질이 응고하기 쉽기 때문이다.
② 종자의 함수량 자체뿐만 아니라 함수량의 변화도 종자의 수명을 단축하는 원인이 된다.
③ 저온 효과는 특히 종자 함수량이 높을 때 크게 나타난다.
④ 콩 종자는 함수량이 많아도 저장온도를 어는점 이하로 내리면 오랫동안 수명을 보존할 수 있다. 그러나 수분함량이
많은 종자를 초저온 하에 두면 결빙으로 인한 해를 받는다. 함수량 적을 시 액화질소(-196℃)에도 안전하다.
⑤ 함수량이 많은 종자를 무산소 상태로 저장하면 혐기성 호흡에 의하여 생성되는 유해물질 때문에 발아가 저해된다.
* 눈의 휴면유도
- 일반적으로 장일조건은 영양생장을 촉진하고 단일조건은 신장생장을 억제하여 휴면아의 형성을 촉진
- 겨울기간 동안 저온에 접촉해야만 휴면이 각성되어 정상적인 생육이 가능
* 2차 휴면 : 발아에 부적당한 환경으로는 고온, 저온, 습윤, 건조, 암흑, 광, 산소 부족 등이 있다.
CO2는 고농도에서는 억제적으로 작용, 저농도에서는 촉진적으로 작용
* 후숙(after ripening) : 많은 종자는 건조에 의해서 어느 수준까지 수분함량이 감소하면 휴면이 소실되는 현상
* 기관의 생장 : 분열 → 확대 → 분화
① 세포 분열단계

② 세포 확대단계
- 산생장설 / 식물세포의 생장에는 액포의 발달 수반, 세포의 생장방향 결정(미세소관의 배열)
* 미세소관은 세포의 생장방향과는 수직으로 배열된다.
③ 세포의 분화단계
- 세포 또는 세포집단이 생리, 생화학적 차이와 함께 구조적인 변화를 가져오는 일련의 과정
- 세포분화의 결과로 다양한 기관형성(배 → 뿌리, 줄기 → 유관속(조직의 원기) → 표피, 피층세포
* 식물의 형태형성
- 광형태 형성 : 빛에 의해 식물의 형태형성이 제어 (빛에 의해 억제 : 줄기신장)
- 암형태형성 : 암상태에서 자란 황백화식물은 줄기가 길고, 떡잎이 겹쳐져 있으며, 엽록소의 축적이 거의 없다.
* 자외선의 파장영역

* 끝눈(정아)이나 곁눈(측아)의 분열조직에서 분화한 엽원기의 생장으로 잎이 형성된다.
* 생장속도(시그모이드곡선)
① 초기(느린시기) : 세포분열에 의한 생장의 준비시기이며, 저장양분에 의존하여 생장하는 시기로 생장속도가 느리다.
② 중기(빠른시기) : 생장체제가 갖추어지게 되면서 세포의 확대생장이 활발하게 이루어지고 대사작용이 왕성해 지면서 급격히 생장하게 된다.
③ 후기(느린시기) : 다시 생장속도가 둔화하는데 이는 광, 수분, 무기양분 등에 대한 경쟁, 생리적 둔화, 생장억제물질의 축적 등의 영향을 받기 때문이다.
* 고온에서는 증산량이 많고 광합성보다 호흡작용이 더 빠르기 때문에 생장속도가 느리다.
야간에 온도가 낮으면 당 함량이 높아지고 뿌리로의 당 이동이 증가하고 호흡에 의한 탄수화물의 소모가 감소하기 때문에 생장에 유리하다.
* 토양산소는 대기 중 산소농도의 1/3이면 뿌리의 생장에 적절한 것으로 알려져 있다. 토성도 생장속도에 영향을 미친다.
일반적으로 사질토양의 경우 생장속도가 빠른 반면에 조직이 치밀하지 못하고 노화가 촉진되는 경향을 보인다.
* 생장상관 - 기타
- 사이토키닌은 잎에 의한 생장억제효과를 감소시켜 겨드랑이눈의 생장을 유도하는 반면,
옥신, 에틸렌, ABA는 겨드랑이 눈의 생장을 억제한다.
- 작물의 재배에서 보상적 상관을 이용하는 것은 적아, 적화, 적과, 적엽, 가지치기 등이다.
* Auxin 농도에 따른 생장 조절

* 브라시노스테로이드 : isoprenoid계열의 화합물로 스콸렌(C30) 을 거쳐 생합성 된다.
* 살리실산의 전구물질 : benzoic acid * 자스몬산의 전구물질 : linoleic aicd
* 지베렐린은 크게 탄소의 수에 따라 탄소 20개와 19개의 두 그룹으로 나눌 수 있으며, 일반적으로 탄소 19개를 가진 지베렐린이 생리활성이 높다.
※ 발아하는 보리 종자에 있어서 GA3에 의한 가수분해 효소 합성은
① 초엽과 배반에서 합성한 GA3 를 배유 쪽으로 방출
② GA3가 호분층으로 확산
③ 호분층에서 α-amylase 등 분해효소 합성, 배유로 분비
④ 전분 등 대분자화합물 분해
⑤ 분해산물이 배반으로 흡수된 후 생장하는 배로 이동
* 옥신의 극성수송
- 옥신의 극성수송은 식물의 줄기-뿌리 극성 발달을 위해서 필수적이다. 식물의 배를 옥신 수송저해제(NPA, TIBA)로 처리하면 줄기와 뿌리 정단에서 심각한 발생 이상과 함께 극성생장 특성이 상실된다. 또한 옥신의 극성수송은 중력의 방향과 무관함. 줄기와 잎에서 옥신의 하향적 극성수송의 주요 통로는 유관속 유조직(주로 물관부)인 것으로 밝혀졌다.
<옥신 극성수송의 화학적 삼투모델>
① IAA는 비해리형(IAAH)으로 수동적으로 또는 음이온형(IAA-)으로 2차 능동공동수송에 의해 세포로 들어옴.
② 세포벽의 pH는 세포막의 H+ -ATP가수분해효소의 활성화에 의해 산성으로 유지됨.
③ 중성 pH를 갖는 세포질에서 음이온형(IAA-)이 우세함.
④ 각 세포의 해부학적 기부 끝에 많이 존재하는 옥신 음이온 유출 운반체를 통하여 IAA-는 세포를 빠져나감.
※ 인돌-3초산의 비해리형(COOH)은 지질 이중층인 세포막을 쉽게 통과한다. 반면, 해리형(COO
-) 옥신은 음전하로 하전 되어 막을 쉽게 통과하지 못한다.
- 성숙한 잎에서 생합성 되는 IAA의 대부분은 체관부를 통하여 식물의 나머지 부위로 비극성 수송되는 것으로 나타난다.
옥신은 체관부 수액 내 다른 성분과 함께 극성수송의 경우보다 훨씬 빠른 속도로 이들 잎에서 식물체의 상하로 이동할 수 있다.
※ 옥신의 대표적인 특성인 극성이동에 따른 발근현상은 대표적인 극성발현의 예이다.
<생장조절물질의 개화조절>
① 옥신 : IAA, NAA 등은 장일식물의 개화를 촉진하나 단일식물의 개화는 억제한다.
② 지베렐린 : GA3 는 저온이나 장일을 대체하여 개화를 유도, 촉진한다.
③ 오이암꽃 착생촉진 : 저온단일, 2,4-D, NAA, 에스렐6
④ 오이수꽃 착생촉진 : 고온장일, 초산은, 지베렐린
※ 시토키닌, 에틸렌 등은 개화를 촉진 MH-30은 개화를 억제하는 것으로 알려져 있다.
* 식물의 노화
- 과실이 성숙할 때에는 과육을 연하게 하는 세포벽 분해효소인 cellulase와 polygalacturonase 등의 합성은 증가
- 잎이 노화할 때는 이들 효소의 생성이 뚜렷하게 관찰되지 않는다.
* 노화의 징후
① 세포 미세구조의 변화 : 일반적으로 엽록체는 잎의 노화가 개시될 때 파괴되는 최초의 세포기관으로 틸라코이드막 구성성분과 스트로마 성분들이 분해된다.
엽록체는 빠르게 분해되는 반면에 핵은 노화의 후기까지 구조적, 기능적으로 완전한 상태로 남아 있다.
② 효소활성 및 유전자 발현의 변화 : 노화의 대표적인 징후는 핵산과 단백질 분해효소가 증가하여 핵산과 단백질의 분해가 가속화되는 것이다.
- 노화가 진행되는 잎에서 감소되는 대표적 단백질은 rubisco로 분해되어 질소원으로 재이용된다.
- 과실이 성숙할 때에는 과육을 연하게 하는 세포벽 분해효소인 cellulase와 polygalacturonase 등의 합성이 증가하나, 잎이 노화할 때에는 이들 효소의 생성이 뚜렷하게 관찰되지 않는다.
※ 잎, 과실의 노화시에 증가되는 가수분해 효소가 상이함.
- 식물의 잎이 노화할 때 특이적으로 증가하는 단백질에는 RNase, proteinase, lipase, alcohol dehydrogenase, methallothionein 및 pathogene-related protein 등이 있다.
- 과실이 노화할 때 특이적으로 발현이 증가하는 유전자들은 에틸렌의 합성에 관여하는 효소, 섬유소 분해효소,
polygalacturonase, glutathionine S-transferase 등이다.
☞ 노화가 진행되면서 발생되는 성숙과 노화에 관련된 단백질은 기관별로 상이하다.
③ 세포막의 변화 : 식물에서 노화가 일어나면서 세포막, 소포체막, 액포막 등으로부터 활성산소의 하나인 과산화물 유리기의 생성이 증가하며 이로 인하여 지방의 과산화반응과 지방산의 탈에스터반응이 일어난다. 노화가 진행되면서 불포화지방산의 산화 중간산물의 양이 크게 증가하며 이와 함께 원형질막의 지질가수분해효소의 양이 급격하게 증가한다.
* 세포의 죽음 과정
① 세포의 죽음으로 예정된 노화과정 : PCD(Programmed Cell Death)
② 괴사(necrosis)로서 냉해, 상처 및 미생물의 침입들과 같이 갑자기 발생하는 외부요인에 의해 일어나는 과정
③ 만성적인 퇴행으로 시간이 경과함에 따라 점차 치명적인 손상이 누적되면서 나타나는 과정 ⇨ 진정한 aging
☞ 손상을 받아 제대로 작용하지 못하는 특정 세포를 식물체 스스로가 선택적으로 제거하기 위해서 다양한 분해효소의
작용에 의하여 세포를 죽게 하는 과정 ⇨ 유전적인 통제를 받는 발달과정의 일환

* 노화와 식물호르몬
에틸렌
에틸렌에 의해 유도되는 일반적인 노화현상으로는 호흡률의 증가, 막투과성의 증가,
그리고 엽록소의 파괴
사이토키닌
세포 내 엽록소, 단백질, DNA, RNA 등의 수준을 유지 또는 증가시켜 노화를 지연시킨다.
뿌리의 활성이 감소하게 되면 사이토키닌의 합성이 줄어들어 잎의 노화가 급격하게 일어난다.
옥 신
잎의 탈리 초기과정을 지연시키는 반면, 탈엽기 또는 과실 성숙 후기에서는 오히려 촉진작용을
나타낸다. 옥신은 엽록소, 핵산, 단백질, 세포막 분해 등의 노화와 관련된 여러 과정들을 변화시킨다.
지베렐린
떡잎, 잎, 열매 등의 조직에서 엽록소의 분해를 억제하고 RNA와 단백질의 분해를 억제하여 식물의
노화를 지연시킨다. 노화가 진행되는 조직에서는 지베렐린 함량이 감소한다.
ABA
노화가 진행되는 동안 단백질분해효소의 합성을 촉진하고 노화관련 단백질의 합성을 촉진한다.
또한 핵산합성을 억제하고 핵산분해효소를 활성화시켜 RNA의 분해를 촉진한다.
※ 콩에서 꽃을 제거하거나 종실이 발달 중인 꼬투리를 제거하면 잎의 노화가 지연된다. 그러나 농업적인 측면에서 본다면 잎의 노화는 작물의 생산성을 떨어뜨리는 요인이 되며 채소작물에서는 수확 후 손실의 큰 요인이 된다.
* 에틸렌 : 발달중인 어린 잎은 완전한 성숙한 잎보다 에틸렌을 더 많이 생성한다.
※ 에틸렌이 옥신 작용에 대한 2차신호 전달자 역할을 하고 있음.
* 시토키닌 생합성 장소(iso - pentenyltransferase)
- 근단 물관부 엽맥, 배주, 미숙종자, 근원기 증축 근관세포, 화서상부, 열매 이층
* 파이토크롬에 의해 유도되는 대표 광가역 반응
- 완두(성체) : 절간신장 저헤
- 도꼬마리(성체) : 개화(광주기 반응) 저해 ⇦ 적색광 효과
※ 적외선(750nm 이상) 중배축의 신장 촉진 특히 군락상태에서 초관 하부에는 원적색광의 비율이 높아 웃자라기 쉽다.
* 잎의 경성운동 : 주야의 주기적인 빛이나 온도변화에 따라 일어나는 식물기관의 경성운동(傾性運動)으로, 보통 수면운동 · 주야운동이라고도 하는데, 예를 들어, 까치콩 · 미모사 · 자귀나무 등의 콩과식물이나 괭이밥과식물의 잎의 취면운동은 잎몸이나 잎자루의 기부에 있는 엽침(葉枕)의 팽압이 빛의 강약에 따라 변화함으로써 일어난다. 이러한 잎의 취면운동을 일으키는 2가지 요인으로는 외위조건(外圍條件)의 변화와 내재리듬이 있다.
* 광주기 감응효과의 지속성 : 일단 유도일장에 감응된 식물은 감응효과가 지속되어 비유도일장 하에서도 꽃눈분화가 계속되어 개화가 가능, 즉 단일식물이 단일처리에 의해 자극을 받고 난 후에는 장일조건에서도 개화가능
- 최소한의 유도일장 처리횟수는 식물의 종과 품종에 따라 다르다. 어떤 식물은 단 한번의 처리로도 꽃눈이 완벽하게 유도되어 영양생장으로 되돌아갈 수 없는가 하면 어떤 식물은 반복처리를 받아야만 감응효과가 지속된다.
* 광주기 자극의 이동 : 서로 다른 두 개체의 줄기 아랫부분을 맞닿게 접목한 후 한쪽에만 단일처리를 하면 단일처리를
하지 않은 다른 쪽의 개체도 개화한다. 광주기의 감응으로 생성된 개화호르몬은 개화자극물질이 정단분열조직으로 이동하여 꽃눈형성을 자극하는데 이 개화호르몬 또는 개화자극물질은 줄기의 위 아래로 이동할 수 있다.
* 광주기성의 작용기구
- 단일식물 : 암기가 길어질 때 연속암기 필요
- 장일식물 : 암기가 짧아질 때 / 광중단 효과에 관계없다.
※ 일장유도 처리된 식물은 비교적 높은 온도에서 개화가 촉진되는 것이 일반적이다.
장일식물에서 암기를 늘려 개화를 억제시킬 경우나 단일식물에서 암기를 늘려 개화를 촉진시킬 경우 모두 온도가 낮으면 개화 억제나 촉진 효과가 감소되며 더욱 긴 유도기간이 필요하게 된다.
즉, 꽃을 재배할 때 밤의 온도에 따라 개화반응이 달라지는 일이 생길 수 있음을 의미한다.
* 광주기 반응의 응용
① 품종간 차이 및 품종의 선발 ② 육종방법에의 이용 ③ 화훼작물의 개화기 조절 ④ 파종기 및 기타 재배상의 문제
☞ 품종간 차이 및 품종의 선발
| ❶ 고위도 지방에서는 감광성이 큰 품종은 일장이 긴 여름까지 영양생장만 계속하다가 그 후 일장이 짧아지면 비로소 유수분화가 시작되므로 가을의 저온으로 완전히 성숙하지 못하므로 적합하지 않고 오히려 감온성이 큰 품종을 재배 ❷ 저위도 지방에서는 감온성이 큰 품종은 영양생장이 충분히 이루어지기 전에 고온이 되므로 일찍 유수를 형성하고 출수하여 다수확을 기대할 수 없다. 따라서 저위도지방에서는 기본영양생장성이 크고 감온성이 적은 품종을 선택 |
* 작물의 발육과 광주기 반응에 끼치는 영향
* 원적색광의 비율(낮은 Pfr : Ptotoal)이 높을 때 식물들은 지상부(줄기신장)로 더 많은 동화산물을 분배한다. 한편 이러한 상관관계는 음지환경에서 자라는 음지식물에서는 뚜렷하지 않다.<양지식물 - 그늘회피 식물>
* 파이토크롬 단백질의 구조와 기능 ; 발색단(색소) + 결손단백질 = 완전단백질
- 고등식물에서 파이토크롬의 발색단은 파이토크로모빌린(phytochoromobilin) 이라고 하는 선형의 테트라피롤(tetrapyrol)
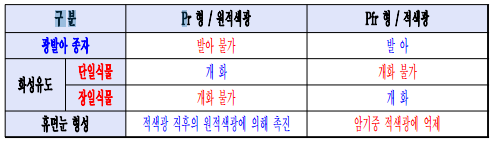
- Pr(cis 이성질체)에 적색광을 조사하면 Pfr(trans 이성질체)로 된다.
* 파이토크롬의 광평형상태
- 적색광 포화조사시 Pr 15% : Pfr 85% / 원적색광 포화조사시 Pr 3% : Pfr 97%로 평형상태 유지
※ Pr형이 적색광을 받으면 대부분의 Pr분자는 Pfr로 바뀐다. 그러나 Pfr의 일부는 적색광도 흡수하여 Pr로 다시 바뀐다.
그러므로 포화조사시에도 상기와 같은 수준의 Pr : Pfr비가 유지가 된다.
* 벼의 조식재배시 수량이 많이 발생 : 기본영양생장성이 긴 감온형
* 작물이 출수 개화에 가장 알맞은 온도와 일장에 놓이더라도 일정한 정도의 기본영양생장을 하지 않으면 출수 개화에 이르지 못하는데 이 성질을 기본영양생장이라고 한다.
※ 혹시 부족하지만 저의 글의 내용이 도움이 조금이라도 되었다면, 단 1초만 투자해 주세요. 로그인이 필요없는 하트♥(공감) 눌러서 블로그 운영에 힘을 부탁드립니다. 그럼 오늘도 좋은 하루 보내세요^^